## 十一、进化
> 原文:[Chapter 11 Evolution](http://greenteapress.com/complexity2/html/thinkcomplexity2012.html)
> 译者:[飞龙](https://github.com/wizardforcel)
> 协议:[CC BY-NC-SA 4.0](http://creativecommons.org/licenses/by-nc-sa/4.0/)
> 自豪地采用[谷歌翻译](https://translate.google.cn/)
生物学乃至整个科学最重要的思想,是通过自然选择的进化论,它声称由于自然选择而创造出新的物种并改变现有的物种。自然选择是个体间遗传差异导致生存和繁殖差异的过程。
在了解生物学的人中,进化论被广泛认为是一个事实,也就是它足以接近事实,如果将来得到纠正,纠正将使中心思想基本保持完整。
尽管如此,许多人并不相信进化论。在皮尤研究中心进行的一项调查中,被调查者被问到,以下哪些断言更贴近他们的观点:
+ 人类和其他生物随时间而进化。
+ 起初,人类和其他生物就以其现在的形式存在。
约有 34% 的美国人选择了第二个(见 <http://www.thearda.com/Archive/Files/Codebooks/RELLAND14_CB.asp>)。
即使在那些认为生物已经进化的人中,只有一半以上的人认为进化的原因是自然选择。 换句话说,只有三分之一的美国人相信进化论是真实的。
这怎么可能? 在我看来,促成因素包括:
+ 有些人认为进化论与他们的宗教信仰之间有冲突。 感觉就像他们不得不拒绝一个,他们拒绝了进化论。
+ 其他人经常被第一组成员积极误导,以至于他们对进化论的许多认识都是错误的。
+ 许多人根本就不了解进化。
对于第一组,我可能没有太多可以做的事,但我认为我可以帮助其他人。 经验上,进化论很难让人理解。 同时,它非常简单:对很多人来说,一旦他们了解进化论,它似乎既明显又无可辩驳。
为了帮助人们从困惑转变为清晰,我找到的最强大的工具就是计算。 我们看到,理论上很难理解的想法,在模拟中出现时很容易理解。 这是本章的目标。
本章的代码位于`chap11.ipynb`中,该书是本书仓库中的 Jupyter 笔记本。使用此代码的更多信息,请参见第?节。
## 11.1 简单的进化
我将从一个简单的模型开始,演示一种基本的进化形式。 根据该理论,以下特征足以产生进化:
+ 复制者:我们需要一批能够以某种方式复制的智能体。 我们将以复制者开始,它们生成它们自己的完美的副本。 稍后我们将添加不完美的副本,即突变。
+ 突变:我们还需要一些种群中的变化,也就是个体之间的差异。
+ 生存和繁殖差异:个体之间的差异必须影响其生存或繁殖的能力。
为了模拟这些特征,我们将定义智能体种群,智能体代表个体。 每个智能体都有遗传信息,称为基因型,这是智能体繁殖时复制的信息。 在我们的模型中 [1],基因型由`N`个二进制数字(零和一)的序列表示,其中`N`是我们选择的参数。
> [1] 模型是主要由 Stuart Kauffman 开发的 NK 模型的变体(参见 <https://en.wikipedia.org/wiki/NK_model>)。
为了产生突变,我们创建了具有多种基因型的种群;稍后我们将探讨创造或增加突变的机制。
最后,为了产生生存和繁殖差异,我们定义了一个函数,将每个基因型映射为一个适应度,其中适应度是一个数量,有关智能体的生存或繁殖能力。
## 11.2 适应性景观
将基因型映射为适应性函数,称为适应性景观。 在景观的隐喻中,每个基因型对应于`N`维空间中的一个位置,并且适应性对应于该位置处的景观的“高度”。对于能够解释这个隐喻的可视化,参见 <https://en.wikipedia.org/wiki/Fitness_landscape>。
在生物学术语中,适应性景观代表一种信息,它是生物体的基因型与其物理形式和能力的关系,后者称为其表现型,以及表现型如何与其环境相互作用。
在现实世界中,适应性景观很复杂,但我们不需要建立现实模型。 为了诱导进化,我们需要基因型和适应性之间的某种关系,但事实证明它可以是任何关系。 为了证明它,我们将使用完全随机的适应性景观。
这是代表适应性景观的类的定义:
```py
class FitnessLandscape:
def __init__(self, N):
self.N = N
self.one_values = np.random.random(N)
self.zero_values = np.random.random(N)
def fitness(self, loc):
fs = np.where(loc, self.one_values,
self.zero_values)
return fs.mean()
```
智能体的基因型,对应其在适应性景观中的位置,由一个 NumPy 的零一数组来表示,称为`loc`。 给定基因型的适应性,是`N`个适应性贡献的平均值,`loc`的每个元素都是一个。
为了计算基因型的适应性,`FitnessLandscape`使用两个数组:`one_values`,其中包含`loc`的每个元素都为 1 时的适应性贡献,以及`zero_values`,其中包含为 0 时的适应度贡献。
`fitness`方法使用`np.where`,如果`loc`中的值为 1,它从`one_values`中选择一个值,如果`loc`中的值为 0,它从`zero_values`中选择一个值。
例如,假设`N=3`:
```py
one_values = [0.1, 0.2, 0.3]
zero_values = [0.4, 0.7, 0.9]
```
这种情况下,`loc = [0, 1, 0]`的适应性是`[0.4, 0.2, 0.9]`的均值,为 0.5。
## 11.3 智能体
接下来我们需要智能体,这是类定义:
```py
class Agent:
def __init__(self, loc, fit_land):
self.loc = loc
self.fit_land = fit_land
self.fitness = fit_land.fitness(self.loc)
def copy(self):
return Agent(self.loc, self.fit_land)
```
智能体的属性是:
`loc`:智能体在适应性景观中的位置。
`fit_land`:`FitnessLandscape`对象的引用。
`fitness`:智能体在`FitnessLandscape`中的适应性,表示为 0 到 1 之间的数字。
`Agent`的这个定义提供了一种简单的`copy`方法,可以精确复制基因型;之后,我们将看到一个带有突变的版本,但突变对于进化来说不是必需的。
## 11.4 模拟
现在我们有了智能体和适应性景观,我将定义一个名为`Simulation`的类,用于模拟智能体的创建,繁殖和死亡。 为了避免陷入困境,我将在这里提供一个简化版本的代码;你可以在本章的笔记本上看到细节。
这是`Simulation`的定义:
```py
class Simulation:
def __init__(self, fit_land, agents):
self.fit_land = fit_land
self.agents = agents
```
`Simulation`的属性是:
+ `fit_land`:`FitnessLandscape`对象的引用。
+ `agents`:`Agent`对象的数组。
`Simulation`中最重要的函数是`step`,它模拟了单个时间步骤:
```py
# class Simulation:
def step(self):
n = len(self.agents)
fits = self.get_fitnesses()
# see who dies
index_dead = self.choose_dead(fits)
num_dead = len(index_dead)
# replace the dead with copies of the living
replacements = self.choose_replacements(num_dead, fits)
self.agents[index_dead] = replacements
```
在每个时间步骤中,一些智能体死亡,一些智能体繁殖。 `step`使用另外三个方法:
+ `get_fitnesses`返回一个数组,包含每个智能体的适应性,按照它们在智能体数组中出现的顺序。
+ `choose_dead`决定哪些智能体在此时间步中死亡,并返回一个数组,包含死亡智能体的索引。
+ `choose_replacements`决定哪些智能体在此时间步中繁殖,在每个智能体上调用`copy`,并返回一个新的`Agent`对象的数组。
在这个版本的模拟中,每个时间步中新智能体的数量等于死亡智能体的数量,所以活动智能体的数量是恒定的。
## 11.5 没有差异
在我们运行模拟之前,我们必须指定`choose_dead`和`choose_replacements`的行为。 我们将从这些函数的简单版本开始,它们不依赖于适应性:
```py
# class Simulation
def choose_dead(self, fits):
n = len(self.agents)
is_dead = np.random.random(n) < 0.1
index_dead = np.nonzero(is_dead)[0]
return index_dead
def choose_replacements(self, n, fits):
agents = np.random.choice(self.agents, size=n, replace=True)
replacements = [agent.copy() for agent in agents]
return replacements
```
在`choose_dead`中,`n`是智能体的数量,`is_dead`是一个布尔数组,对于此时间步骤内死亡的智能体为`True`。 在这个版本中,每个智能体都有相同的死亡概率:0.1。 `choose_dead`使用`np.nonzero`来查找`is_dead`的非零元素的索引(`True`被视为非零)。
在`choose_replacements`中,`n`是在此时间步骤中复制的智能体数量。 它使用`np.random.choice`带替换地选择`n`个智能体。 然后它在每个上调用`copy`,并返回一个新的`Agent`对象列表。
这些方法不依赖于适应性,所以这种模拟没有生存或繁殖差异。 因此,我们不应期待看到进化。 但是,我们怎么辨别呢?
## 11.6 进化的证据
进化的最具包容性的定义是,种群中基因型分布的变化。 进化是一种聚合效应:换句话说,个体不会进化;但种群会。
在这个模拟中,基因型是高维空间中的位置,因此很难将其分布中的变化可视化。 但是,如果基因型改变,我们预计它们的适应性也会改变。 所以我们将将适应性分布的变化用作进化的证据。 具体来说,我们将看看种群中适应性的均值和标准差。
在我们运行模拟之前,我们必须添加一个`Instrument`,它是在每个时间步骤后更新的对象,计算一个感兴趣的统计量,并将结果存储在一个序列中,我们稍后可以绘制它。
这是所有仪器的父类:
```py
class Instrument:
def __init__(self):
self.metrics = []
```
下面是`MeanFitness`的定义,`MeanFitness`是一个仪器,计算每个时间步的种群平均适应性:
```py
class MeanFitness(Instrument):
def update(self, sim):
mean = np.nanmean(sim.get_fitnesses())
self.metrics.append(mean)
```
现在我们准备好运行模拟了。 为了最小化起始种群中随机变化的影响,我们使用同一组智能体启动每个模拟。 为了确保我们探索整个适应性景观,我们由每个位置的一个智能体开始。 以下是创建模拟的代码:
```py
N = 8
fit_land = FitnessLandscape(N)
agents = make_all_agents(fit_land, Agent)
sim = Simulation(fit_land, agents)
```
`make_all_agents`为每个位置创建一个智能体; 本章的实现在笔记本中。
现在我们可以创建并添加`MeanFitness`仪器,运行模拟并绘制结果:
```py
instrument = MeanFitness()
sim.add_instrument(instrument)
sim.run()
sim.plot(0)
```
模拟维护了`Instrument`对象列表。 在每个时间步之后,它在列表中的每个仪器上调用`update`。
模拟运行后,我们使用`Simulation.plot`绘制结果,它接受索引作为参数,使用索引从列表中选择一个`Instrument`并绘制结果。 在这个例子中,只有一个`Instrument`,索引为 0。
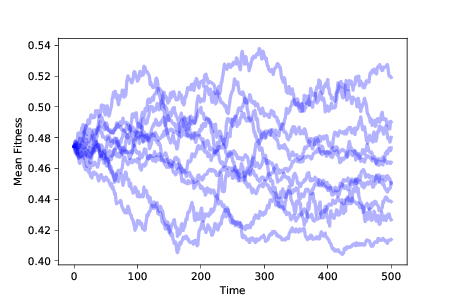
图 11.1:随着时间的推移,10 次模拟的平均适应性,没有生存或繁殖差异
图?显示了运行这个模拟 10 次的结果。 种群的平均适应性随机移动。 由于适应性的分布随时间变化,我们推断表现型的分布也在变化。 按照最具包容性的定义,这种随机游走是一种进化。 但它不是一个特别有趣的类型。
特别是,这种进化并不能解释生物物种如何随时间变化,或者如何出现新的物种。 进化论是强大的,因为它解释了我们在自然界看到的似乎无法解释的现象:
+ 适应性:物种与其环境的相互作用似乎太复杂,太巧妙,并且偶然发生。 自然系统的许多特征看起来好像是设计出来的。
+ 增加的多样性:地球上的物种数量随时间而普遍增加(尽管有几个时期的大规模灭绝)。
+ 增加的复杂性:地球上的生命史起始于相对简单的生命形式,后来在地质记录中出现了更复杂的生物体。
这些是我们想要解释的现象。 到目前为止,我们的模型并没有完成这个任务。
## 11.7 生存差异
让我们再添加一种成分,生存差异。 以下是继承`Simulation`并覆盖`choose_dead`的类的定义:
```py
class SimWithDiffSurvival(Simulation):
def choose_dead(self, fits):
n = len(self.agents)
is_dead = np.random.random(n) > fits
index_dead = np.nonzero(is_dead)[0]
return index_dead
```
现在生存的概率取决于适应性;事实上,在这个版本中,智能体在每个时间步骤中幸存的概率是其适应性。
由于适应性低的智能体更有可能死亡,因此适应性高的智能体更有可能生存足够长的时间来繁殖。 我们预计适应性低的智能体的数量会随时间而减少,适应性高的智能体的数量会增加。
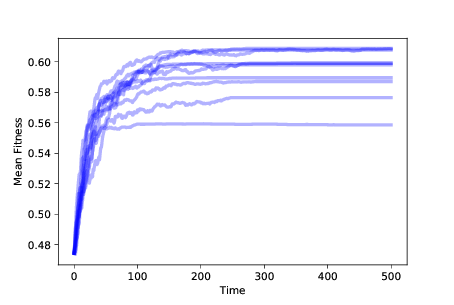
图 11.2:随着时间的推移,10 次模拟中的适应性均值,带有生存差异
图?显示了随着时间的推移,10 次模拟中的适应性均值,带有生存差异。 平均适应性起初会迅速增加,但会逐渐平稳。
你或许可以弄清楚为什么它会平稳:如果在特定位置只有一个智能体并且它死了,它就会使这个位置变空。没有突变,就没有办法让它再次被占领。
在`N = 8`的情况下,该模拟以 256 个智能体开始,它们占用了所有可能位置。 占用位置的数量随时间而减少;如果模拟运行时间足够长,最终所有智能体将占用相同的位置。
所以这个模拟开始解释适应性:增加的适应性意味着,物种在它的环境中生存得更好。 但是占用位置的数量随时间而减少,所以这个模型根本无法解释增加的多样性。
在本章的笔记本中,你将看到差异化繁殖的效果。 正如你所预料的那样,差异化繁殖也会增加平均适应性。但没有突变,我们仍然没有看到增加的多样性。
## 11.8 突变
在目前的模拟中,我们以可能的最大多样性开始 - 在景观的每个位置都有一个智能体 - 并以可能的最小多样性结束,所有智能体都在一个位置。
这与自然界发生的情况几乎相反,它显然以单个物种开始,这种物种随时间而分化为今天的地球上数百万甚至数十亿物种(见 <https://en.wikipedia.org/wiki/Global_biodiversity>)。
使用我们模型的完美复制,我们从未看到增加的多样性。 但是如果我们加上突变,再加上生存和繁殖差异,我们距离理解自然界的进化就更近了一步。
以下是继承`Agent`并覆盖`copy`的类定义:
```py
class Mutant(Agent):
prob_mutate = 0.05
def copy(self):
if np.random.random() > self.prob_mutate:
loc = self.loc.copy()
else:
direction = np.random.randint(self.fit_land.N)
loc = self.mutate(direction)
return Mutant(loc, self.fit_land)
```
在这种突变模型中,每次我们调用`copy`时,都有 5% 的突变机会。 在突变的情况下,我们从当前位置选择一个随机方向 - 即基因型中的一个随机位 - 并翻转它。 这是`mutate`:
```py
def mutate(self, direction):
new_loc = self.loc.copy()
new_loc[direction] ^= 1
return new_loc
```
运算符`^=`计算“异或”;操作数 1 具有翻转一位的效果(请参阅 <https://en.wikipedia.org/wiki/Exclusive_or#Bitwise_operation>)。
现在我们有了突变,我们不必在每个位置都放置一个智能体。 相反,我们可以以最小变化开始:所有智能体在同一位置。
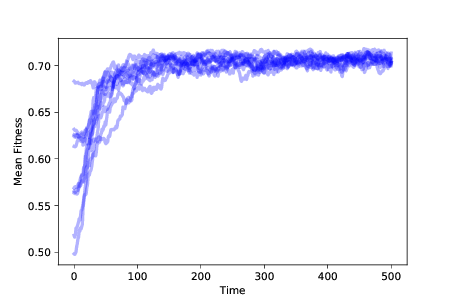
图 11.3:随着时间的推移,10 次模拟中的适应性均值,带有突变、生存繁殖差异
图?显示了 10 次模拟的结果,带有突变和生存繁殖差异。 在任何情况下,种群都会向最大适应性的位置进化。
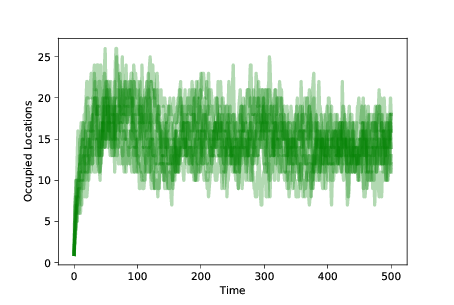
图 11.4:随着时间的推移,10 次模拟的占用位置的数量,带有突变和生存繁殖差异。
为了测量种群的多样性,我们可以绘制每个时间步后占用位置的数量。 图?展示了结果。 我们以同一地点的 100 个智能体开始。 随着突变的发生,占用位置的数量迅速增加。
当智能体发现适应性高的位置时,它更有可能生存和繁殖。 适应性较低的位置上的智能体最终消失。 种群在整个景观中随时间而移动,直到大多数智能体处于适合性最高的位置。
此时,系统达到平衡,突变以相同的速率占据新的位置,生存差异导致适合性低的位置清空。
平衡中的占用位置的数量,取决于突变率和生存差异的程度。 在这些模拟中,任何点处的独特占用位置的数量通常为 10-20。
重要的是要记住,这个模型中的智能体不会移动,就像生物体的基因型没有改变一样。 当智能体死亡时,它可能会留下一个空位。 当发生突变时,它可以占据一个新的位置。 当智能体从某些地方消失并出现在其他地方时,种群会在景观中移动,就像生命游戏中的滑翔机一样。 但生命体不会进化;但种群可以。
## 11.9 物种形成
进化论说,自然选择改变了现有的物种并创造了新的物种。 在我们的模型中,我们看到了变化,但我们并没有看到新的物种。 在模型中,还不清楚新物种是什么样。
在有性繁殖的物种中,如果两种生物能够繁殖并产生丰富的后代,则被视为同一物种。 但是模型中的智能体不会再现性行为,所以这个定义不适用。
在无性繁殖的生物中,如细菌,物种的定义并不明确。 一般来说,如果一个种群的基因型形成一个簇,那么它就被认为是一个物种,也就是说,如果种群内的遗传差异比种群间的差异小。
在我们可以对新物种建模之前,我们需要能够识别景观中的智能体簇,这意味着我们需要定义位置之间的距离。 由于位置是用二进制数字串表示的,因此我们将距离定义为基因型中不同的位数。 `FitnessLandscape`提供了`distance `方法:
```py
# class FitnessLandscape
def distance(self, loc1, loc2):
return np.sum(np.logical_xor(loc1, loc2))
```
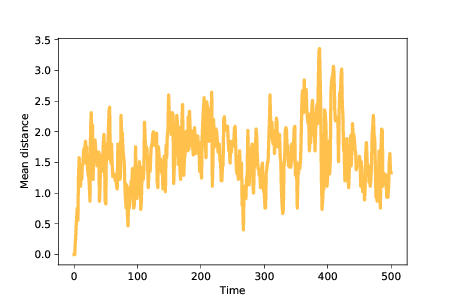
图 11.5:智能体随时间变化的平均距离
`logical_xor`函数计算“异或”,不同的元素为`True`,相同的元素为`False`。
为了量化种群的分散,我们可以计算每对智能体之间距离的平均值。 在本章的笔记本中,你会看到`MeanDistance`仪器,它会在每个时间步骤后计算这个度量。
图? 展示了智能体随时间的平均距离。 因为我们从相同的突变开始,所以初始距离为 0。随着突变的发生,平均距离增加,在种群遍布景观时达到最大值。
一旦智能体发现最佳位置,平均距离就会减小,直到种群达到平衡,由于突变引起的距离增加通过距离的减小而平衡,因为远离最佳位置的智能体更有可能死亡。 在这些模拟中,平衡时的平均距离接近 1.5;也就是说,大多数智能体距离最佳位置只有 1-2 个突变。
现在我们准备寻找新的物种。 为了模拟一种简单的物种形成,假设一个种群在不变的环境中演化,直到它达到稳定状态(就像我们在自然界发现的一些物种,似乎在很长一段时间内变化很小)。
现在假设我们改变环境,或者将种群转移到新的环境中。 一些旧环境中适应性较高的特性,可能会在新环境中适应性较低,反之亦然。
我们可以通过运行模拟来模拟这些情景,直到种群达到稳定状态,然后改变适应性景观,然后恢复模拟,直到种群再次达到稳定状态。
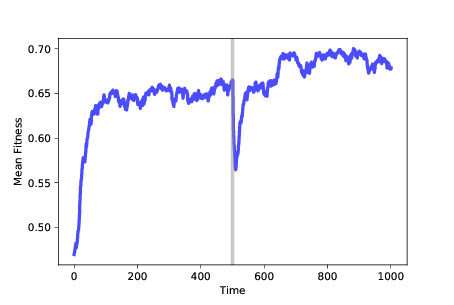
图 11.6:随时间变化的适应性均值。在 500 步之后,我们改变了适应性景观
图?展示了这样的模拟结果。 再次,我们从随机位置开始,使用 100 个相同的突变体,并运行 500 个时间步骤的模拟。 在这个时候,许多智能体处于最佳位置,在这个例子中,其适应性接近 0.65。 智能体的基因型形成一个簇,智能体之间的平均距离接近 1。
经过 500 步之后,我们运行`FitnessLandscape.set_values`,这改变了适应性景观; 然后我们恢复模拟。 平均适应性会立即下降,因为旧景观中的最佳位置并不比新景观中的随机位置好。
然而,当种群迁移到新景观时,平均适应性会迅速增加,最终会找到新的最佳值,其适应度接近 0.75(在这个例子中恰好更高,但不一定是)。
一旦种群达到稳定状态,它就会形成一个新的簇,智能体之间的平均距离再次接近 1。
现在,如果我们计算智能体之前和之后的位置之间的距离,平均而言,它们相差超过 6。 簇之间的距离远大于每个簇内的距离,因此我们可以将这些簇解释为不同的物种。
## 11.10 总结
我们已经看到,突变以及生存和繁殖差异,足以导致适应性的增加,多样性的增加,并产生简单形式的物种。 这种模型并不是现实的;自然系统中的进化要比这复杂得多。 相反,它意味着一个“充足定理”;也就是说,模型的特征足以产生我们试图解释的行为(参见 <https://en.wikipedia.org/wiki/Necessity_and_sufficiency>)。
从逻辑上讲,这个“定理”并不能证明,自然界中的进化仅仅由这些机制引起,但是由于这些机制确实以多种形式出现在生物系统中,所以认为它们至少有助于自然进化,是合理的。
同样,该模型并不能证明这些机制总是会导致进化。 但是我们在这里看到的结果是可靠的:在几乎所有包含这些特征的模型中 - 不完美的复制者,变异性和繁殖差异 - 发生了进化。
我希望这一观察有助于揭开进化的神秘面纱。 当我们观察自然系统时,进化看起来很复杂。 而且由于我们主要看到了进化的结果,而没有看到这个过程,所以难以想象和相信。
但在模拟中,我们看到整个过程,而不仅仅是结果。 通过包含最少的一系列特征来产生进化 - 暂时忽略了生物生命的巨大复杂性 - 我们可以将进化看作是一个令人惊讶的简单,不可避免的想法。
## 11.11 练习
本章的代码位于本书仓库的 Jupyter 笔记本`chap11.ipynb`中。 打开笔记本,阅读代码并运行单元格。 笔记本包含本章的练习。 我的解决方案在`chap11soln.ipynb`中。
